Жаропонижающие средства для детей назначаются педиатром. Но бывают ситуации неотложной помощи при лихорадке, когда ребенку нужно дать лекарство немедленно. Тогда родители берут на себя ответственность и применяют жаропонижающие препараты. Что разрешено давать детям грудного возраста? Чем можно сбить температуру у детей постарше? Какие лекарства самые безопасные?
Как правило, именно белки ответственны за функциональную активность мембран.
К таким белкам относятся разнообразные ферменты, транспортные белки, рецепторы, каналы, белки, образующие поры (аквапорины), то есть разнообразные белковые структуры, которые обеспечивают уникальность функций каждой мембраны.
Мембранные белки по биологической роли можно разделить на три группы:
I – белки-ферменты, обладающие каталитической активностью,
II – рецепторные белки, специфически связывающие те или иные вещества,
III – структурные белки.
Белки-ферменты
Наиболее распространены среди всех мембранных белков. В их число входят как интегральные (мембранные АТФазы), так и периферические (ацетилхолинэстераза, кислая и щелочная фосфатазы, РНКаза) белки.
Ферменты – большие молекулы, в то время как размеры молекул веществ (субстратов), вступающих в ферментативные реакции, обычно в тысячи раз меньше. Фермент взаимодействует с субстратом небольшим участком своей поверхности – активным центром. Специфичность фермента всегда определяется тем, насколько поверхность его активного центра соответствует поверхности субстрата. Этот принцип структурного соответствия повсеместно используется и в работе белков клеточных мембран. В дополнение к этому надо учесть, что конформация внедряющихся в мембрану белков зависит от мембранного бислоя, так что и их ферментативная активность контролируется мембранными липидами. Этот контроль может реализоваться благодаря как влиянию на сродство к субстратам или на их доступность, так и воздействию на длительность жизни (прочность) белковых ассоциатов мембранных ферментов, образующихся в клеточной мембране.
Ферменты входят в состав как плазматических, так и внутриклеточных мембран. Например, на наружной мембране эпителиальных клеток, выстилающих пищеварительные органы, имеются ферменты, осуществляющие расщепление питательных веществ еще до того, как они попадут внутрь клетки (этот процесс, открытый отечественным физиологом А.М. Уголевым носит название «мембранное пищеварение»). Наружная мембрана клеток печени содержит более 20 различных ферментов.
Мембранные ферменты нуждаются в контакте с окружающими их липидами. Когда их извлекают из липидного окружения (например, когда липиды экстрагируются из мембраны неполярными растворителями), работа мембранных ферментов нарушается (меняются особенности кинетики или характера влияния посторонних веществ или же вовсе прекращается). Активность таких мембранных ферментов удается частично восстановить, если к ним добавить липидные мицеллы.
Анализ природы липидов, активирующих мембранные ферменты, демонстрирует отсутствие строгой специфичности - определяющим является гидрофильно-липофильный коэффициент липидной смеси. В ряде случаев активировать делипидированный фермент удается даже детергентом. Однако такой реактивированный фермент теряет способность воспринимать регулирующие сигналы извне, которые управляли его работой в «живой» мембране.
Активирующее действие липидов на мембранные ферменты может быть, по меньшей мере, двояким. Во-первых, в присутствии липидов может меняться форма молекулы мембранного фермента, так что его активный центр становится доступным для субстрата. Во-вторых, липиды могут играть роль организатора ансамбля или конвейера, состоящего из многих ферментов.
Молекулы мембранных ферментов содержат большие неполярные гидрофобные участки. Поэтому в водной среде они агрегируют, из-за чего большая часть активных центров маскируется. В присутствии липидов мембранные ферменты организуются в ансамбли, окруженные аннулярными липидными молекулами, и их ферментативная активность может проявиться в полной мере. Для нормальной работы мембранных ферментов существенно, чтобы окружающие их липиды находились в жидком агрегатном состоянии.
Рецепторные белки
Рецепторными называют белки, специфически связывающие те или иные низкомолекулярные вещества. При связывании специфических лигандов рецепторные белки обратимо меняют свою форму. Эти изменения запускают внутри клетки ответные химические реакции. Таким способом клетка воспринимает различные сигналы, поступающие из внешней среды, и отвечает на них .
Белки-рецепторы и белки, определяющие иммунную реакцию клетки, – антигены, также могут быть как интегральными, так и периферическими компонентами мембраны.
Часто рецепторы входят в состав более сложных мембранных комплексов, содержащих белки-исполнители. Например, холинорецептор воспринимает сигнал от нейромедиатора и передает его на белок-каналообразователь. Эта реакция открывает проницаемость мембраны для ионов натрия и калия и формирует возбуждающий потенциал.
: характеристика и структурные принципы
1. Структура мембранных белков
Основная роль липидов в составе мембран заключается в стабилизации бислойной структуры, а белки являются активными компонентами биомембран. Мы обсудим некоторые принципы, оказавшиеся полезными для выяснения структурных особенностей мембранных белков. Мы приведем примеры, иллюстрирующие эти принципы.
На заре развития мембранологии полагали, что мембранные белки по своей структуре довольно гомогенны и уложены в виде 3-слоев по поверхности бислоя. Сейчас скорее склонны считать, что по крайней мере у трансмембранных белков те их участки, которые погружены в мембрану, содержат а-спирали. Конечно, очень хотелось бы сделать какие-то однозначные выводы по этому поводу, но они должны основываться на фактических данных. Перед лицом огромного структурного разнообразия растворимых белков приходишь к заключению, что интегральные мембранные белки могут оказаться гораздо сложнее, чем мы сейчас представляем. Классификация растворимых белков по типам структур была проведена только после того, как установили с высоким разрешением структуру более 100 различных белков. Что касается трансмембранных белков, то это удалось сделать только в одном случае - для белка фотосинтетического реакционного центра бактерий. Вместе с электронно-микроскопическими данными низкого разрешения о структуре бактериородопсина это единственный источник, на котором может основываться построение моделей для большинства других трансмембранных белков.
Еще один важный момент - способы прикрепления белков к мембране. Они схематически представлены на рис. 3.1.
1. Связывание с белками, погруженными в бислой. В качестве примеров можно привести Fi-часть Н + -АТРазы, которая связывает ся с Fo-частью, погруженной в мембрану; можно упомянуть также некоторые белки цитоскелета.
2. Связывание с поверхностью бислоя. Это взаимодействие имеет в первую очередь электростатическую природу или гидрофобную. На поверхности некоторых мембранных белков имеются гидрофобные домены, образующиеся благодаря особеностям вторичной или третичной структуры. Указанные поверхностные взаимодействия могут использоваться как дополнение к другим взаимодействиям, например к трансмембранному заякориванию.
3.Связывание с помощью гидрофобного «якоря»; эта структура обычно выявляется как последовательность неполярных аминокислотных остатков. Некоторые мембранные белки используют в качестве якоря кова-лентно связанные с ними жирные кислоты или фосфолипиды.
4.Трансмембранные белки. Одни из них пересекают мембрану только один раз, другие - несколько раз.
Различиями между наружными и внутренними мембранными белками не задается однозначно способ их прикрепления к бисЛою; эти различия определяют лишь относительную силу их связывания.
2. Очистка мембранных белков
Для очистки интегральных мембранных белков и получения их в биохимически активной форме необходимы детергенты, позволяющие солюбилизировать белки и сохранить их в растворе. Соответствующие требования к детергентам и правилам обращения с ними создают дополнительные проблемы помимо тех, с которыми обычно сталкиваются при очистке белков. Для выделения интегральных мембранных белков разработано много специальных методов, однако большинство схем очистки основано на тех же хроматографических и гидродинамических методиках, которые используются для растворимых белков. Это хроматография на ДЭАЭ-целлюлозе, сефарозе или гидроксила-патите, гель-фильтрация, центрифугирование в градиенте плотности сахарозы и т. д. Очень важен правильный выбор детергента, поскольку именно детергент разрушает биомембрану, занимая место липидов, окружающих тот или иной белок, и определяет стабильность белка в растворе. Механизмы действия детергентов рассмотрены в обзоре.
2.1. ДЕТЕРГЕНТЫ
В течение последних двух десятилетий появилось очень много детергентов, пригодных для очистки интегральных мембранных белков. В принципе нужно пытаться найти такой детергент, который не нарушал бы вторичную и третичную структуры мембранных белков, а лишь замещал бы большинство или все мембранные липиды, контактирующие с гидрофобными участками белковой молекулы. Конечной целью солюбилизации является встраивание белка в детергентиую мицеллу; последующая стратегия очистки состоит в разделении таких белково-детергентных комплексов.
Первая проблема - это подбор оптимальных условий солюбили-зации изучаемого белка. Детергенты, денатурирующие белки, не подходят для решения такой деликатной задачи. С другой стороны, многие детергенты недостаточно эффективно разрушают мембраны и образуют белоксодержащие смешанные мицеллы. Такие детергенты могут быть либо слишком гидрофобными, либо слишком гидрофильными для эффективного смешивания с мембранными липидами и - при достаточно высокой их концентрации - для превращения бислоя в глобулярные смешанные мицеллы. Сначала надеялись, что выбор необходимого детергента удастся систематизировать с помощью одного параметра, называемого гидро-фильно-липофильным балансом. Этот параметр, изменяющийся от 1 до 20, используется при получении сурфактантов в качестве меры относительной гидрофобности. Действительно, получены некие корреляции, из которых следует, что значение ГЛБ детергента может использоваться для предсказания его поведения в биологических системах. Вообще говоря, можно сказать, что детергенты со значением ГЛБ в диапазоне от 12,5 до 14,5 являются наиболее эффективными растворителями интегральных мембранных белков. Однако впоследствии выяснилось, что поиск оптимальных детергентов для определенного мембранного белка требует учета многих факторов и всегда должен сопровождаться эмпирической проверкой. Необходимо учитывать следующее.
1.Максимальная солюбилизация исследуемого белка. Критерием является переход белка в супернатант после центрифугирования, при котором происходит осаждение мембраны.
2.Солюбилизация белка в нужной форме. Обычно речь идет о сохранении его ферментативной активности, но иногда используются определенные спектральные характеристики или наличие конкретных белковых ассоциатов. Кроме того, необходимым условием является стабильность белка после солюбилизации. В некоторых случаях для поддержания биохимической активности вместе с детергентом добавляют экзогенные фосфолипиды. В качестве примера можно привести получение лактозопермеазы Е. coliи белка натриевого канала. Иногда для стабилизации белка после солюбилизации добавляют глицерол или другой полиол. Имеет смысл использовать также ингибиторы протеаз и проводить солюбилизацию в условиях, сводящих к минимуму вероятность их протеолитического расщепления.
3.Возможность использования детергента в данной методике. Необходимо прежде всего учитывать заряд детергента, поведение при данном значении рН, ККМ и размер мицелл детергента. Последние свойства особенно важны. Детергенты с низкой ККМ, образующие крупные мицеллы, не удаляются при диализе или ультрафильтрации из-за слишком низкой концентрации мономеров детергента. С практической точки зрения это означает, что если концентрировать белок с помощью ультрафильтрации, то будет возрастать и концентрация детергента с низкой ККМ, а это может привести к денатурации белка. По этой причине многие исследователи предпочитают использовать детергенты с высокими ККМ, например октилглюкозид, соли желчных кислот или более современные цвиттерионные детергенты. Весьма ценными являются полистиреновые смолы, такие, как биобидз SM-2. Они избирательно связываются с детергентами типа тритон Х-100, удаляют их из раствора и позволяют обойтись вообще без диализа. Еще один фактор, который необходимо учитывать, - это поглощение света детергентом. Некоторые детергенты, например тритон Х-100, поглощают в ближней УФ-области, что делает невозможным определение концентрации белка по измерению оптической плотности при длине волны 280 нм.
С учетом всех этих факторов становится понятно, почему во многих случаях при выделении интегральных мембранных белков приходится использовать разные детергенты. Например, для солюбилизации можно применять тритон Х-100, а разделение с помощью ДЭАЭ-целлюлозы лучше проводить в присутствии октилглюкозида. Детергенты можно менять на стадии хроматографии, во время центрифугирования в градиенте плотности, а в некоторых случаях - с помощью диализа. Следует иметь в виду, что детергент, непригодный для солюбилизации определенного белка, может быть очень эффективным для сохранения белка в растворе после замены детергента. Очистку почти всегда следует проводить при избытке детергента в растворе, в противном случае равновесие будет сдвинуто в сторону агрегации мембранных белков, а не в сторону образования белково-детергентных комплексов. В некоторых случаях подобная агрегация может быть даже желательна, и последняя стадия очистки может состоять в удалении детергента. Но, как правило, при недостатке детергента происходят необратимое осаждение и потеря белка.
Необходимость поддержания концентрации детергента на определенном уровне создает дополнительнее трудности помимо тех, с которыми обычно сталкиваются при очистке белков; о некоторых из них мы уже говорили. Проблемы возникают и при использовании стандартного метода высаливания при высокой концентрации сульфата аммония: во многих случаях белок осаждается в комплексе с детергентом и липидом. Поскольку солевой раствор имеет высокую плотность, а детергент в агрегате - относительно низкую, то при центрифугировании преципитат будет оставаться на поверхности. Важно помнить, что очистке подвергаются белково-детергентные комплексы, нередко со значительным количеством связанного фосфолипида. Это сказывается на качестве разделения при хроматографировании, а также на результатах характеристики конечного прорастворимых белков, нужно определить число и молекулярную массу полипептидных субъединиц, их стехиометрию, размер и, возможно, форму молекулы, а также, если это необходимо, биохимическую активность.
Детергент 1 В табл. 1 и 2 перечислены наиболее широко используемые детергенты и указаны их свойства, важные для обсуждаемых нами вопросов. Эмпирически наиболее эффективными являются: 1) неионные детергенты (тритон Х-100, октилглюкозид); 2) соли желчных кислот (холат, дезоксихолат); 3) цвиттерионные детергенты (CHAPS, цвнттергент). Но выбор детергента, наиболее приемлемого для солюбилизации и очистки определенного мембранного фермента, по-прежнему осуществляется методом проб и ошибок. |
ККМ, мМ | Мол.масса | 1 Размер А мнцеллы | .грегационис число | х Удельный объем, мл/г | Ссылки |
| Долецилсульфат | 1,33 | 288 | 24 500 | 85 | 0,864 | |
| натрия | ||||||
| Холат натрия " | 3 | 408 | 2100 | 5 | 0,778 | (612, 1383] |
| Дезоксихолат | 0,91 | 392 | 23 000 | 55 | 0,771 | |
| натрия " | ||||||
| 0,11 | 538 | 68 000 | 12 | 0,973 | ||
| Тритои Х-100 2) | 0,24 | 628 | 90 000 | 140 | 0,908 | |
| Твин 80 2) | 0,012 | 1300 | 76 000 | 60 | 0,8% | |
| Лаурилдиметил- | 2,2 | 229 | 17 000 | 75 | 1,112 | |612] |
| аминоксид | ||||||
| ^-D-Октил- | 25 | 293 | 8000 | 27 | 0,820 | |
| ^-D-Лаурил- | 0,16 | 510 | 50 000 | 98 | 0,820 | |
| мальтозид | ||||||
| CHAPS | 8 | 615 | 6150 | 10 | 0,802 | |
| Цвиттергеит | 3,6 | 335 | - | - | 0,957 |
3. ХАРАКТЕРИСТИКА ОЧИЩЕННЫХ ИНТЕГРАЛЬНЫХ МЕМБРАННЫХ БЕЛКОВ
Характеристика очищенных мембранных белков, даже самых простых, может составлять определенные трудности. Как и в случае
3.1 МОЛЕКУЛЯРНАЯ МАССА СУБЪЕДИНИЦ
Электрофорез в полиакриламидном геле в присутствии додецил-сульфата натрия - это обычная методика, но в случае интегральных мембранных белков при ее применении возникают особые проблемы. В этом методе додецилсульфат связывается с полипептидными цепями, и комплексы белок-ДНС разделяются в полиакриламидном геле в соответствии с их стоксовыми радиусами, которые в большинстве случаев зависят от молекулярной массы. Молекулярную массу определяют, сравнивая электрофоретическую подвижность данного комплекса и известного стандарта. Однако связывание ДСН с неизвестным белком может качественно отличаться от связывания со стандартами, и тогда будет получен неправильный результат. Подобная ситуация наблюдается для интегральных мембранных белков с высоким содержанием неполярных аминокислотных остатков. С большинством растворимых белков ДСН образует комплексы в соотношении 1,4 г ДСН на 1 г белка, а с белками, содержащими большой процент неполярных остатков, может связываться больше детергента. Возникающий при этом дополнительный отрицательный заряд приводит к аномальному повышению электрофоретической подвижности, и определяемая молекулярная масса оказывается меньше, чем на самом деле. Возможна и другая ситуация. Связывающийся с ДСН мембранный белок может находиться в не полностью развернутом состоянии, что тоже приведет к аномальному повышению электрофоретической подвижности из-за образования более компактного комплекса белок-ДСН. Все эти эффекты весьма существенны. Например, лактозопермеаза имеет кажущуюся мол. массу 33 ООО, если измерять ее с помощью электрофореза в ПААГ в присутствии ДСН; в действительности же, как показывают результаты генетического анализа, ее мол. масса равна 46 ООО. Во многих случаях удается оценить молекулярную массу более точно, если построить график Фергюсона, представляющий собой зависимость электрофоретической подвижности от содержания акриламида как для стандартных белков, так и для исследуемого белка. Этот график зависит от радиуса Стокса и в меньшей степени - от заряда комплекса. Например, по результатам электрофореза в 12%-ном акриламидном геле одна из субъединиц цитохромно- г о комплекса Е. coliимеет кажущуюся мол. массу 28 ООО, а из графика Фергюсона получается величина 43 ООО, что совпадает с мол. массой, рассчитанной по данным о секвенировании соответствующей ДНК.
Еще одна проблема - возможное наличие четвертичной структуры. Некоторые мембранные белки агрегируют даже в присутствии ДСН. Например, гликофорин А или белок оболочки бактериофага М13 при электрофорезе в полиакриламидных гелях с ДСН находятся в основном в виде димеров. Иногда агрегация еще более усиливается при нагревании смеси белок-ДСН. Такая картина наблюдается, например, для субъединиц как митохондри-альной, так и бактериальной терминальных оксидаз. Чтобы оценить способность белка к необратимой агрегации, следует провести сравнительный анализ результатов электрофореза в полиакрила-мидном геле с ДСН для прогретых и непрогретых проб. Сходная проблема иногда возникает из-за присутствия детергента, использованного при очистке мембранного белка. Этот детергент необходимо удалить и заменить на ДСН, поскольку в некоторых случаях наблюдается четкая зависимость электрофоретической подвижности от присутствия детергента, с помощью которого солюбилизнровали фермент.
Итак, есть основания думать, что оценка молекулярной массы субъединиц сильно неполярных интегральных мембранных белков, определенная с помощью электрофореза в ПААГ с ДСН, может оказаться неверной. К несчастью, простая альтернатива этому методу отсутствует, и правильную величину часто получают либо по данным о полной первичной последовательности, либо с помощью точного гидродинамического анализа.
3.2 ОПРЕДЕЛЕНИЕ МОЛЕКУЛЯРНОЙ МАССЫ НАТИВНОГО БЕЛКА С ПОМОЩЬЮ ГИДРОДИНАМИЧЕСКИХ МЕТОДОВ
Применение этих методов для мембранных белков может быть сопряжено с большими трудностями, вызванными связыванием детергента. Чтобы оценить это в полной мере, рассмотрим вначале простой растворимый белок, для которого установлена мол. масса субъединиц с помощью электрофореза в ПААГ с ДСН и необходимо узнать, чем он является в неденатурированной, активной форме - мономером, димером или олигомером более высокого порядка. Для определения молекулярной массы белков часто используется гель-фильтрация, включающая сравнение со стандартными белками; здесь возникают проблемы, связанные с тем, что все стандартные белки имеют глобулярную форму, а исследуемый белок может быть не глобулярным, а слегка удлиненным. Такой белок с мол. массой 50 000 может элюировать со скоростью, соответствующей мол. мае
се 100 ООО. В связи с этим колонка для гель-фильтрации должна быть прокалибрована в соответствии со значениями радиуса Стокса, т. е. с размерами «эквивалентной гидродинамической сферы», а кроме того, параллельно необходимо использовать какой-либо другой метод. Обычно измеряют скорость седиментации с помощью либо аналитического ультрацентрнфугирования, либо центрифугирования в градиенте плотности сахарозы. Коэффициент седиментации равен
![]()
где м - молекулярная масса белка,
v - его парциальный удельный объем, ij - вязкость раствора, б - плотность раствора.
Поскольку е и Ч известны, aRcможно определить с помощью гель-фильтрации, остаются только две неизвестные величины - v и м. Для водорастворимых белков v можно вычислить исходя из аминокислотного состава или непосредственно измерить либо просто принять равным 0,72-0,75 мл/г. Таким образом, измерив S 0 , можно найти м.
Рассмотрим теперь ситуацию с мембранным белком. Здесь возникают дополнительные проблемы, поскольку гидродинамическая частица - это белково-детергентный комплекс, поэтому м и v в данном случае являются молекулярной массой и удельным объемом комплекса, М к и К,. К сожалению, К, нельзя оценить, не зная ничего о составе комплекса. В этом случае для нахождения молекулярной массы белка используют два метода.
1.Прямо измеряют количество связанного детергента на 1 г белка. Для этого используют спектральные методы или радиоактивно меченный детергент, а для выделения комплексов применяют различные методы, например гель-фильтрацию. Установив относительное содержание белка и детергента в комплексе, значение К, получают как средневзвешенное соответствующих величин для чистого белка и чистого детергента. После этого без труда находят м„ а поскольку соотношение между белком и детергентом в комплексе известно, находят молекулярную массу белка.
2.Измеряют S 0 в средах с разными значениями плотности раствора д. Такие среды обычно получают, используя смеси НгО и D2O. Из графика зависимости S° от qнаходят как Л/„ так и v t . При этом предполагается, что К, - это средневзвешенное соответствующих величин для чистого белка и чистого детергента.
ОцеНИВ Квело* и взяв детергент из таблиц, получают молекулярную массу белковой составляющей м,.
Для построения графика зависимости 5° от qпроводят аналитическое центрифугирование. Можно проводить центрифугирование и в градиенте плотности сахарозы, используя смеси Н2О и D2O, но анализ результатов в этом случае гораздо сложнее, хотя принципиально не отличается от предыдущего случая.
Альтернативный способ определения молекулярной массы нативной формы мембранного белка состоит в равновесном ультрацентрифугировании. Распределение вещества в состоянии равновесия таково, что наклон графика зависимости логарифма концентрации от г 2 равен

где г - расстояние от центра ротора до данной точки в центрифужной пробирке, W- частота вращения.
Если величина У известна или ее легко оценить, как для большинства растворимых белков, эта задача решается достаточно просто. Что касается мембранных белков, то в этом случае определяют на-
Таблица 3. Связывание детергентов с некоторыми мембранными белками
клон указанной прямой при разных значениях q, получаемых смешиванием НгО и D2O. Как и ранее, одновременно находят М к и К, и далее определяют молекулярную массу белка.
Если в комплексе присутствует третий компонент, возникают дополнительные проблемы. В любом случае все описанные процедуры весьма сложны и могут давать ошибочные результаты. Количество детергента, связанного с очищенными интегральными мембранными белками, может быть весьма существенным - от 0,3 до 1,5 от массы белка, и даже небольшие ошибки в этой величине приведут к значительному искажению молекулярной массы белка. В табл. 3.3 приведены данные о количестве детергентов, присутствующих в некоторых белковых препаратах. Заметим, что растворимые белки с этими детергентами не связываются; это опять свидетельствует о том, что за связывание с детергентом ответственна именно неполярная часть белка, обычно контактирующая с мембранными липидами.
3.3 МЕТОД РАДИАЦИОННОЙ ИНАКТИВАЦИИ
Метод радиационной инактивации для определения размера мишени все чаще применяется при исследовании мембранных белков. Изучать можно как очищенные белки, так и неочищенные препараты, в том числе интактные биомембраны. Суть метода состоит в определении доли белковых молекул, получающих повреждения при облучении. Для этого используют ферментативные методы связывания гормонов или других лигандов или спектральные методы. Процедура состоит в следующем. Образец, обычно замороженный, подвергают высокоэнергетическому облучению. Через разные промежутки времени отбирают пробы, размораживают их и проводят измерения. Повреждения белка под действием излучения выявляют, например, с помощью электрофореза в ПААГ с ДСН. Как показывает опыт, некоторые субъединицы полностью утрачивают биологическую активность при внесении радиационного повреждения в любое место полипептидной цепи. Ключевым моментом является то, что, чем крупнее белковая молекула, тем больше вероятность ее повреждения и, следовательно, вероятность инактивации. Эта вероятность зависит не от формы молекулы, а от ее массы. Обычно для того, чтобы облегчить интерпретацию результатов, параллельно облучают белок с известной молекулярной массой. Если исследуемый белок содержит более одной субъединицы, возникают определенные трудности при анализе результатов. Повреждение одной субъединицы не обязательно сопровождается разрывом ковалентных связей в других субъединицах. Поэтому для ферментов, состоящих из разных субъединиц, обладающих неодинаковыми активностями, могут быть получены разные размеры мишени в зависимости от метода определения степени инактивации.
Примечательной особенностью метода является то, что его можно использовать для изучения интегральных мембранных белков insitu. Возникающие при этом артефакты и проблемы рассмотрены в работе. Одна из очевидных проблем - необходимость использования высокоэнергетического излучения. В связи с этим большинство работ приходится проводить в сотрудничестве с лабораториями, в которых имеются соответствующие источники и освоены специальные методы анализа.
3.4 СПЕКТРАЛЬНЫЕ МЕТОДЫ И ВТОРИЧНАЯ СТРУКТУРА
Для определения содержания а-спиралей и /3-слоев в мембранных белках используют несколько методов. В отсутствие трехмерной организации на их основе можно попытаться построить соответствующие модели. Чаще всего используется метод кругового дихроизма. Все более широкое применение находят инфракрасная и рамановская спектроскопия, а также ЯМР.
1. Метод кругового дихроизма основан на измерении разности поглощения лево- и правополяризованного света; эта оптическая активность является мерой хиральности молекул, или мерой их асимметрии. В дальней ультрафиолетовой области КД определяется в основном поглощением амидов карбонильных групп полипептидного остова. При наличии участков вторичной структуры, например а-спиралей, спектр КД имеет вполне определенные особенности, связанные с особенностями электронного окружения амидиых групп в этих структурах. Анализизуя спектр КД белков, его обычно представляют как сумму компоиеитов, отвечающих поглощению разных участков белковой молекулы: а-спиралей, /3-слоев и случайных клубков. Определив тем или иным способом спектры каждой из этих структур, производят их суммирование, подбирая соответствующие коэффициенты таким образом, чтобы было достигнуто наилучшее соответствие измеренному спектру. Подобранные весовые коэффициенты представляют собой ту долю, которая приходится в молекуле на каждый из типов вторичной структуры.
Эти методы были разработаны для растворимых белков, но нет никаких оснований сомневаться, что их можно с успехом применять и для мембранных белков. Скорее всего у последних имеются участки с такими же типами вторичной структуры, как и у растворимых белков, и при их изучении возникнут такие же трудности. Некоторые белки можно изучать insitu, используя суспензии мембран. Примерами такого рода являются бактериородопсин из пурпурной мембраны Halobacteriumhalobiumи Са 2 + -АТРаза из мембраны саркоплазмати-ческого ретикулума. Очищенные мембранные белки можно исследовать с помощью КД и в присутствии детергентов, если поглощение последних в дальней УФ-области не слишком велико, или в составе реконструированных везикул. Здесь возникают две проблемы: 1) дифференциальное светорассеяние, когда размер мембранных частиц гораздо больше длины волны света; 2) выравнивание поглощения из-за концентрирования белка в мембранах или везикулах, т. е. из-за негомогенности его распределения в растворе. Эти артефакты могут быть весьма существенными, однако их можно учесть с помощью соответствующих методов.
К сожалению, для внутренних мембранных белков отсутствуют структурные данные высокого разрешения, поэтому точная интерпретация спектров КД невозможна. За исключением нескольких случаев, разные спектральные методы не использовались для изучения одного и того же белка и количественное сравнение результатов не проводилось. Интересно, что для бактериородопсина, который исследовали методами КД, ИК и ЯМР, во всех трех случаях были получены одинаковые результаты, свидетельствующие о значительном содержании в этом белке 3-слоев. Тем не менее у каждого метода имеются существенные недостатки. Так, данные о высоком содержании в бактериородопсине Д-слоев в значительной мере зависят от способа учета оптических артефактов. Судя по данным электронно-микроскопической реконструкции, харатеризующимся относительно низким разрешением, в бактериородопсине 80% приходится на долю а-спира-лей, а 0-слои отсутствуют совсем. Чтобы понять причину этих несоответствий, необходимо провести структурный анализ белка с атомным разрешением. Имеются еще два белка, пронизывающие мембрану, с высоким содержанием, и а-токсин Staphylococcusaureus. Оба этих белка участвуют в образовании пор в бислое.
2. Инфракрасная спектроскопия и спектроскопия комбинационного рассеяния. Эти методы не только позволяют получить сведения о конформации мембранных липидов, но и могут использоваться для исследования вторичной структуры белков. Колебательный спектр полипептидного остова зависит от типа вторичной структуры и дает информацию о содержании в молекуле а- и /3-структур. Этими методами можно исследовать высушенные на воздухе пленки, водные суспензии мембран, а также очищенные белки как в присутствии детергента, так и в составе реконструированных везикул. Например, по данным ИК-спектроскопии с преобразованием Фурье комплекс Са 2 + -АТРазы в мембране состоит в основном из а-спиральных участков и участков, имеющих кон-формацию статистического клубка, а гидрофобный белок миелин в реконструированных везикулах имеет как а-, так и /3-участки.
3. ЯМР-спектроскопия также может использоваться для изучения мембранных белков. Однако возможности метода в этом случае ограничены, что связано главным образом с относительно медленными движениями интегральных мембранных белков insitu и в комплексах с детергентом. Поэтому такой мощный метод, как двумерный ЯМР, который может дать детальную картину конформацион-ного состояния сравнительно небольших белков в растворе, пока непригоден для изучения мембранных белков. Более приемлем метод ЯМР твердых образцов. Большими возможностями обладают методы 2 Н- и |3 С-ЯМР, хотя до сих пор они применялись не очень широко. Получены данные об усредненной конформации остова и динамике боковых цепей. Следует отметить, что методы ЯМР твердого состояния не только не используются широко, но в большинстве случаев их и нельзя использовать. Тем не менее в тех редких ситуациях, когда их применение оказывается возможным, они являются очень ценными.
3.5 ФЕРМЕНТАТИВНАЯ АКТИВНОСТЬ
Одним из наиболее важных методов характеристики очищенных мембранных белков несомненно является определение биохимической активности. При этом используются в основном такие же критерии, как и для растворимых белков, но могут возникать и свои трудности. Первая из них связана с тем, что биохимическая активность мембранных белков часто очень сильно зависит от связывания с белком липидов и детергентов. Потеря активности может быть как обратимой, так и необратимой. Целесообразно иметь какую-то оценку удельной активности исследуемого белка invivo или в составе мембран до солюбилизации. Избыток детергента может оказывать ингибирующий эффект, например за счет разбавления неполярных субстратов в популяции мицелл и уменьшения ферментативной активности. Измеряя активность любого мембранного белка, необходимо иметь в виду, что insitu он находится в окружении липидов, обеспечивающих оптимальную активность. Вторая проблема связана с белками, обладающими «трансбислойной» активностью; примерами могут служить белки, образующие каналы, и транспортные белки. В этих случаях необходимо учитывать перемещение растворенных веществ из одного компартмента в другой.
3.6 ЧЕТВЕРТИЧНАЯ СТРУКТУРА И ХИМИЧЕСКОЕ СШИВАНИЕ
Многие мембранные ферменты представляют собой комплексы, состоящие из нескольких субъединиц. В качестве примера можно привести Н + -АТРазу, Na + /К + -АТРазу, митохондриальные комплексы электронного транспорта и фотосинтетические реакционные центры. Некоторые интегральные мембранные белки прочно связаны с растворимыми белками с помощью нековалентных взаимодействий. В Е. coliFo-компонент, содержащий по данным электрофореза в ПААГ-ДСН три типа субъединиц, образует протонный канал, aFi, состоящий из пяти типов субъединиц, содержит активный центр, участвующий в гидролизе АТР. Для таких белков очень важно определить характер субъединиц, стехиометрию комплекса и ближайшие взаимодействия его компонентов. Это весьма непростая задача даже тогда, когда белковый комплекс уже изолирован. Возникающие здесь проблемы по существу не отличаются от таковых для растворимых белковых комплексов, но имеются и свои дополнительные сложности.
Прежде всего следует иметь в виду, что взаимодействие между субъеднницами очень сильно зависит от типа липидов и детергентов, с которыми связаны белки. Например, сукцинатде-гидрогеназа Е. coliпри солюбилизации ее с помощью луброла РХ представляется состоящей из четырех субъединиц, а при солюбилизации большинством других детергентов, в том числе тритоном Х-100, - только из двух. Известно, что оперон sdhкодирует все четыре полипептида, а форма из двух субъединиц имеет аномальный спектр ЭПР. Таким образом, ясно, что invivo фермент состоит из четырех субъединиц. Однако сукцинатдегидрогеназной активностью обладают обе формы, поэтому используемые биохимические критерии важны для заключения, была ли солюбилизирована правильная форма.
Еще одна проблема связана с тем, что в бислое мембранные белки могут образовывать комплексы из-за высокой их локальной концентрации. При солюбилизации же независимо от используемого детергента может произойти разбавление мембранных белков и их разъединение. По закону действующих масс это приведет к диссоциации комплексов, в которых взаимодействие между компонентами не очень сильное. Часто бывает трудно определить, какой комплекс образуется insitu, а какой - прн солюбилизации и очистке. Подобные проблемы возникают при исследовании многих сложных систем, например системы /3-адренергетический рецептор-адеиилатци-клаза, цепи электронного транспорта у митохондрий, системы мик-Росомных цитохромов Р450 и b$.
Для изучения стехиометрии субъединиц и их ассоциации в очищенном комплексе используется всего несколько методов: 1) химичес кое сшивание; 2) количественный анализ N-концевых аминокислот; 3) определение отношения массы субъединиц в ДСН-полиакриламидных гелях путем определения интенсивности окрашивания, с помощью радиоавтографии или иммуноблоттинга. Каждый метод имеет свои ограничения, но все они использовались на практике. Например, стехиометрию пяти субъединиц никотинового ацетил-холииового рецептора определяли с помощью количественного анализа N-коицевых аминокислот, а трех субъедиииц Fo-компоиента Н + -АТРазы Е. coli- с помощью разделения в ДСН-полиакриламидных гелях. Заметим, что кумасси бриллиантовый синий, обычно используемый для окрашивания белков после разделения в ПААГ-ДСН, связывается предпочтительнее с белками, содержащими основные аминокислотные остатки, и существуют примеры сильно неполярных внутренних мембранных белков, которые лишь едва окрашиваются.
Химическое сшивание применялось для определения ближних взаимодействий как в очищенных белковых комплексах, так и в комплексах insitu. Для анализа ближних взаимодействий в мембранных белках используется несколько специфических гидрофобных сшивающих агентов. Некоторые из них представлены в табл. 3.4. Применяемые методы не отличаются от таковых для растворимых систем. Продукты сшивания обычно анализируют с помощью электрофореза в ПААГ, часто с использованием расщепляемых сшивающих агентов, что позволяет анализировать полипептиды. Применяют также антитела к индивидуальным полипептидам для иммуноблоттинга после электрофореза в ПААГ-ДСН, чтобы идентифицировать компоненты каждого из образовавшихся продуктов. Можно было бы предположить, что при относительно большом времени жизни реагентов белки в биомембранах будут сшиваться в результате простой диффузии в бислое. Однако, по данным нескольких работ, это не так: продукты сшивания представляют собой специфические белковые ассоциаты, а не случайные образования. Так, в фотосинтетической мембране Rhodobactercapsulataобразуются сшивки лишь между субъединицами компонентов реакционного центра, а также между реакционным центром и «антенным» комплексом В870, участвующим в передаче энергии реакционному центру.
И наконец, отметим, что химическое сшивание часто использовалось для идентификации интегральных мембранных белков, которые связываются с известными растворимыми компонентами. В качестве примера можно привести сшивание 1) а- и b-субъединиц Fo-kom-понента Н + -АТРазы с /3-субъединицей растворимого компонента Fi; 2) цитохрома с с субъединицами митохондриальной цитохром с-оксидазы; 3) пептидных гормонов с рецепторами гормонов.
Таблица 4. Некоторые сшивающие реагенты, использовавшиеся для определения четвертичной структуры мембранных белков "
Будущее медицины - персонифицированные методы избирательного воздействия на отдельные системы клетки, которые ответственны за развитие и течение конкретного заболевания. Основным классом терапевтических мишеней при этом являются мембранные белки клетки как структуры, ответственные за обеспечение непосредственной передачи сигналов в клетку. Уже сегодня почти половина лекарств воздействуют именно на клеточные мембраны, и дальше их будет только больше. Знакомству с биологической ролью мембранных белков посвящена данная статья.
Структура и функции клеточной мембраны
Из школьного курса многие помнят устройство структурной единицы организма - клетки. Особое место в устройстве живой клетки играет плазмалемма (мембрана), которая отделяет внутриклеточное пространство от окружающей ее среды. Таким образом, главная ее функция - создание барьера между клеточным содержимым и внеклеточным пространством. Но это не единственная функция плазмолеммы. Среди других функций мембраны, связанных в первую очередь с мембранными белками, выделяют:
- Защитную (связывание антигенов и предупреждение их проникновения в клетку).
- Транспортную (обеспечение обмена веществ между клеткой и средой).
- Сигнальную (встроенные рецепторные белковые комплексы обеспечивают раздражимость клетки и ее ответ на различные воздействия извне).
- Энергетическую - преобразование разных форм энергий: механической (жгутики и реснички), электрической (нервный импульс) и химической (синтез молекул аденозинтрифосфорной кислоты).
- Контактную (обеспечение связи между клетками при помощи десмосом и плазмодесм, а также складок и выростов плазмолеммы).

Строение мембран
Мембрана клетки - это двойной слой липидов. Бислой образуется благодаря наличию в молекуле липидов двух частей с разными свойствами - гидрофильного и гидрофобного участка. Наружный слой мембран образован полярными «головками» с гидрофильными свойствами, а гидрофобные «хвосты» липидов обращены внутрь бислоя. Кроме липидов, в структуру мембран входят белки. В 1972 году американские микробиологи С.Д. Сингер (S. Jonathan Singer) и Г.Л. Николсон (Garth L. Nicolson) предложили жидкостно-мозаичную модель строения мембраны, согласно которой, мембранные белки «плавают» в бислое липидов. Эта модель была дополнена немецким биологом Каем Зимонсом (1997) в части образования определенных, более плотных участков с ассоциированными белками (липидных рафтов), которые свободно дрейфуют в бислое мембраны.
Пространственная структура мембранных белков
В различных клетках соотношение липидов и белков различно (от 25 до 75% белков в пересчете на сухую массу), и расположены они неравномерно. По расположению белки могут быть:
- Интегральными (трансмембранными) - встроенными в мембрану. При этом они пронизывают мембрану, иногда неоднократно. Их внеклеточные участки часто несут цепи олигосахаридов, формируя гликопротеиновые кластеры.
- Периферическими - расположены преимущественно на внутренней стороне мембран. Связь с липидами мембраны обеспечивается за счет водородных обратимых связей.
- Заякоренными - преимущественно расположены с наружной стороны клетки и «якорем», удерживающим их на поверхности, является молекула липида, погруженная в бислой.

Функционал и обязанности
Биологическая роль мембранных белков многообразна и зависит от их структуры и расположения. Среди них выделяют рецепторные белки, канальные (ионные и порины), транспортеры, моторы и структурные белковые кластеры. Все виды мембранных белков-рецепторов в ответ на какое-либо воздействие меняют свою пространственную структуру и формируют ответ клетки. Например, рецептор инсулина регулирует поступление глюкозы внутрь клетки, а родопсин в чувствительных клетках органа зрения запускает каскад реакций, что приводят к возникновению нервного импульса. Роль мембранных белков-каналов заключается в транспорте ионов и поддержании разницы их между внутренней и внешней средой. Например, натрий-калиевые насосы обеспечивают обмен соответствующих ионов и веществ. Порины - сквозные белки - участвуют в переносе молекул воды, транспортеры - в переносе некоторых веществ против градиента концентраций. У бактерий и простейших движение жгутиков обеспечивают молекулярные белковые моторы. Структурные мембранные белки поддерживают саму мембрану и обеспечивают взаимодействие других белков плазмолеммы.
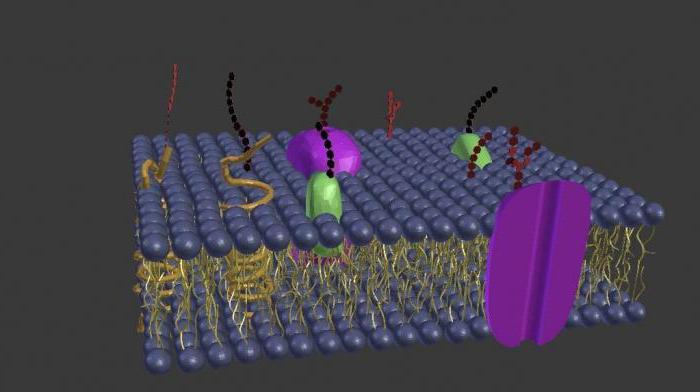
Белки для мембраны, мембрана для белков
Мембрана - это динамическая и очень активная среда, а не инертная матрица для белков, которые в ней расположены и работают. Она существенно влияет на работу мембранных белков, а липидные рафты, перемещаясь, формируют новые ассоциативные связи белковых молекул. Многие белки просто не работают без партнеров, и межмолекулярное их взаимодействие обеспечивается характером липидного слоя мембран, структурная организация которого, в свою очередь, зависит от структурных белков. Нарушения в этом тонком механизме взаимодействия и взаимозависимости приводят к нарушению функций мембранных белков и целому ряду заболеваний, таких как диабет и злокачественные опухоли.
Структурная организация
Современные представления о структуре и строении мембранных белков основаны на том, что в мембранной периферической части большинство из них состоит редко из одной, чаще из нескольких ассоциированных олигомеризующихся альфа-спиралей. Причем именно такая структура является залогом выполнения функции. Однако именно классификация белков по типам структур может принести еще немало сюрпризов. Более чем из ста описанных белков наиболее изученным по типу олигомеризации мембранным белком является гликофорин А (белок эритроцитов). Для трансмембранных белков ситуация выглядит сложнее - описан лишь один белок (фотосинтетический реакционный центр бактерий - бактериородопсин). Учитывая высокую молекулярную массу мембранных белков (10-240 тысяч дальтон), у молекулярных биологов широкое поле для исследований.
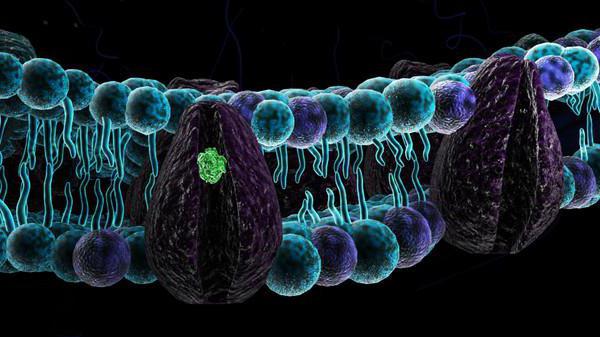
Сигнальные системы клетки
Среди всех белков плазмолеммы особое место принадлежит рецепторным белкам. Именно они регулируют, какие сигналы поступят в клетку, а какие нет. У всех многоклеточных и некоторых бактерий передача информации осуществляется посредством специальных молекул (сигнальных). Среди этих сигнальных агентов выделяют специально секретируемые клетками), небелковые образования и отдельные ионы. Последние могут выделяться при повреждении соседних клеток и запускать каскад реакций в виде болевого синдрома, главного защитного механизма организма.
Мишени для фармакологии
Именно мембранные белки являются главными мишенями применения фармакологии, так как именно они и есть те точки, через которые идет большинство сигналов. «Нацелить» лекарственный препарат, обеспечить его высокую селективность - вот главная задача при создании фармакологического средства. Избирательное воздействие только на конкретный тип или даже подтип рецептора - это влияние только на один тип клеток организма. Такое селективное воздействие может, например, отличить опухолевые клетки от нормальных.

Лекарства будущего
Свойства и особенности мембранных белков уже сегодня используются в создании лекарств нового поколения. Эти технологии основаны на создании модульных фармакологических структур из нескольких молекул или наночастиц, «сшитых» друг с другом. «Нацеливающая» часть узнает на мембране клетки определенные рецепторные белки (например, связанные с развитием онкологических заболеваний). К этой части добавляется разрушающий мембрану агент или блокатор процессов производства белков в клетке. Развивающийся апоптоз (программа собственной гибели) или другой механизм каскада внутриклеточных превращений приводит к желаемому результату воздействия фармакологического средства. В результате мы имеем лекарство с минимумом побочных эффектов. Первые такие лекарства по борьбе с раком уже проходят клинические испытания и вскоре станут залогом высокоэффективной терапии.

Структурная геномика
Современная наука о белковых молекулах все интенсивнее переходит на информационные технологии. Экстенсивный путь исследований - изучить и описать все, что можно, сохранить данные в компьютерных базах и потом искать пути применения данных знаний - такова цель современных молекулярных биологов. Всего лишь пятнадцать лет назад стартовал глобальный проект «геном человека», и мы уже имеем секвенированную карту генов человека. Второй проект, цель которого - определить пространственное строение всех «ключевых белков», - структурная геномика - пока далек от завершения. Пространственная структура определена пока только для 60 тысяч более чем из пяти миллионов белков человека. И пусть пока ученые вырастили лишь светящихся поросят и холодоустойчивые помидоры с геном лосося, технологии структурной геномики остаются этапом научного познания, практическое приложение которого не заставит долго себя ждать.
К мембранным белкам относятся белки, которые встроены в клеточную мембрану или мембрану клеточной органеллы или ассоциированы с таковой. Около 25 % всех белков являются мембранными.
Биохимическая классификация
По биохимической классификации мембранные белки делятся наинтегральные и периферические .
- Интегральные мембранные белки прочно встроены в мембрану и могут быть извлечены из липидного окружения только с помощью детергентовили неполярных растворителей. По отношению к липидному бислою интегральные белки могут быть трансмембранными политопическими или интегральными монотопическими.
- Периферические мембранные белки являются монотопическими белками. Они либо связаны слабыми связями с липидной мембраной, либо ассоциируют с интегральными белками за счёт гидрофобных, электростатических или других нековалентных сил. Таким образом, в отличие от интегральных белков они диссоциируют от мембраны при обработке соответствующим водным раствором (например, с низким или высоким pH, с высокой концентрацией соли или под действием хаотропного агента). Эта диссоциация не требует разрушения мембраны.
Мембранные белки могут быть встроены в мембрану за счёт жирнокислотных или пренильных остатков либогликозилфосфатидилинозитола, присоединённых к белку в процессе их посттрансляционной модификации.
Еще один важный момент - способы прикрепления белков к мембране:
1. Связывание с белками, погруженными в бислой. В качестве примеров можно привести F1-часть Н + - АТРазы, которая связывается с Fo-частью, погруженной в мембрану; можно упомянуть также некоторые белки цитоскелета.
2. Связывание с поверхностью бислоя. Это взаимодействие имеет в первую очередь электростатическую природу (например, основный белок миелина) или гидрофобную (например, поверхностно-активные пептиды и, возможно, фосфолипазы). На поверхности некоторых мембранных белков имеются гидрофобные домены, образующиеся благодаря особенностям вторичной или третичной структуры. Указанные поверхностные взаимодействия могут использоваться как дополнение к другим взаимодействиям, например к трансмембранному заякориванию.
3. Связывание с помощью гидрофобного "якоря"; эта структура обычно выявляется как последовательность неполярных аминокислотных остатков (например, у цитохрома 65). Некоторые мембранные белки используют в качестве якоря ковалентно связанные с ними жирные кислоты или фосфолипиды.
4. Трансмембранные белки. Одни из них пересекают мембрану только один раз (например, гликофорин), другие - несколько раз (например, лактозопермеаза; бактериородопсин).
Мембранные липиды
Мембранные липиды - это амфипатические молекулы, самопроизвольно формирующие бислои. Липиды нерастворимы в воде, однако легко растворяются в органических растворителях. В большинстве животных клеток они составляют около 5О% массы плазматической мембраны. В участке липидного бислоя размером 1 х 1 мкм находится приблизительно 5 х 1ОО тыс. молекул липидов. Следовательно плазматическая мембрана небольшой животной клетки содержит примерно 1О липидных молекул. В клеточной мембране присутствуют липиды трех главных типов:
1) фосфолипиды (наиболее распространенный тип);сложные липиды, содержащие глицерин, жирные кислоты, фосфорную кислоту и азотистое соединение.
Типичная молекула фосфолипида имеет полярную голову и два гидрофобных углеводородных хвоста. Длина хвостов варьирует от 14 до 24 атомов углерода в цепи. Один из хвостов содержит, как правило, одну или более цис-двойных связей (ненасыщенный углеводород), тогда как у другого (насыщенный углеводород) двойных связей нет. Каждая двойная связь вызывает появление изгиба в хвосте. Подобные различия в длине хвостов и насыщенности углеводородных цепей важны, поскольку они влияют на текучесть мембраны.
Амфипатические молекулы, находящиеся в водном окружении, обычно агрегируют, при этом гидрофобные хвосты оказываются спрятанными, а гидрофильные головы остаются в контакте с молекулами воды. Агрегация такого типа осуществляется двумя способами: либо путем образования сферических мицелл с хвостами обращеными внутрь, либо путем формирования бимолекулярных пленок, или бислоев, в которых гидрофобные хвосты располагаются между двумя слоями гидрофильных голов.
Два основных фосфолипида, которые присутствуют в плазме - это фосфатидилхолин (лецитин) и сфингомиелин. Синтез фосфолипидов происходит почти во всех тканях, но главным источником фосфолипидов плазмы служит печень. Тонкий кишечник также поставляет в плазму фосфолипиды, а именно лецитин, в составе хиломикрон. Большая часть фосфолипидов, которые попадают в тонкий кишечник (в том числе и в виде комплексов с желчными кислотами), подвергается предварительному гидролизу панкреатической липазой. Этим обьясняется, почему полиненасыщенный лецитин, добавленный в пищу, влияет на содержание линолеата в фосфолипидах плазмы не больше, чем триглицериды кукурузного масла в эквивалентных количествах.
Фосфолипиды являются неотьемлемым компонентом всех клеточных мембран. Между плазмой и эритроцитами постоянно происходит обмен фосфатидилхолином и сфингомиелином. Оба эти фосфолипида присутствуют в плазме в качестве составных компонентов липопротеинов, где они играют ключевую роль, поддерживая в растворимом состоянии неполярные липиды, такие как триглицериды и эфиры холестерина. Это свойство отражает амфипатический характер молекул фосфолипидов - неполярные цепи жирных кислот способны взаимодействовать с липидным окружением, а полярные головы - с водным окружением (Jackson R.L. ea, 1974).
2) Холестерол. Холестерин - это стерин, содержащий стероидное ядро из четырех колец и гидроксильную группу.
Это соединение обнаруживается в организме как в виде свободного стерина, так и в форме сложного эфира с одной из длинноцепочечных жирных кислот. Свободный холестерин - компонент всех клеточных мембран и та основная форма, в которой холестерин присутствует в большинстве тканей. Исключение представляют кора надпочечников, плазма и атероматозные бляшки, где преобладают эфиры холестерина. Кроме того, значительная часть холестерина в кишечной лимфе и в печени тоже этерифицирована.
Холестерин содержится в составе липопротеин ов либо в свободной форме, либо в виде эфиров с длинноцепочечными жирными кислотами. Он синтезируется во многих тканях из ацетил-CoA и выводится из организма желчь ю в виде свободного холестерола или солей желчных кислот. Холестерол является предшественником других стероид ов, а именно кортикостероидов, половых гормонов, желчных кислот и витамина D . Он является соединением, типичным для метаболизма животных, и содержится значительных количествах в продуктах животного происхождения: яичном желтке, мясе, печени и мозге.
Плазматические мембраны эукариот содержат довольно большое количество холестерола - приблизительно одну молекулу на каждую молекулу фосфолипида. Помимо регулирования текучести холестерол увеличивает механическую прочность бислоя. Молекулы холестерола ориентируются в бислое таким образом, чтобы их гидроксильные группы примыкали к полярным головам фосфолипидных молекул
3) гликолипиды
Гликолипиды - это липидные молекулы, принадлежащие к классу олигосахаридсодержащих липидов, которые обнаруживаются только в наружной половине бислоя, а их сахарные группы ориентированы к поверхности клетки.
Гликолипиды это сфинголипиды, у которых к NH группе сфингазина присоединен остаток ЖК, а к кислороду сфингазина присоединены следующие группы: олигосахаридные цепи, Gal, Glc, GalNAc (нейраминовая кислота) – ганглиозиды. Gal или Glc – цереброзиды. сульфосахара Glc-SO3H, Gal-SO3H – сульфолипиды.
Гликолипиды обнаруживаются на поверхности всех плазматических мембран, однако их функция неизвестна. Гликолипиды составляют 5% липидных молекул наружного монослоя и сильно различаются у разных видов и даже в разных тканях одного вида. В животных клетках они синтезируются из сфингозина - длинного аминоспирта - и называются гликосфинголипидами.
Структура их в целом аналогична структуре фосфолипидов, образованных из глицерола. Все гликолипидные молекулы различаются по числу сахарных остатков в их полярных головах. Один из простейших гликолипидов – галактоцереброзид
Липидам в составе мембран отводят, в первую очередь, структурные свойства - они создают бислой, или матрикс, в котором размещаются активные компоненты мембраны - белки. Именно белки придают разнообразным мембранам уникальность и обеспечивают специфические свойства. Многочисленные мембранные белки выполняют следующие основные функции: обусловливают перенос веществ через мембраны (транспортные функции), осуществляют катализ, обеспечивают процессы фото- и окислительного фосфорилирования, репликацию ДНК, трансляцию и модификацию белков, рецепцию сигналов и передачу нервного импульса и др.
Принято делить мембранные белки на 2 группы: интегральные (внутренние) и периферические (наружные). Критерием такого разделения служит степень прочности связывания белка с мембраной и, соответственно, степень жесткости обработки, необходимой для извлечения белка из мембраны. Так, периферические белки могут высвобождаться в раствор уже при промывке мембран буферными смесями с низкой ионной силой, низкими значениями рН в присутствии хелатирующих веществ, например этилендиаминотетраацетата (ЭДТА), связывающих двухвалентные катионы. Периферические белки выделяются из мембран при таких мягких условиях, поскольку связаны с головками липидов или с другими белками мембраны при помощи слабых электростатических взаимодействий, либо с помощью гидро-фобных взаимодействий - с хвостами липидов. Наоборот, интегральные белки представляют собой амфифильные молекулы, имеют на своей поверхности большие гидрофобные участки и располагаются внутри мембраны, поэтому для их извлечения требуется разрушить бислой. Для этих целей наиболее часто используют детергенты или органические растворители. Способы прикрепления белков к мембране довольно разнообразны (рис. 4.8).
Транспортные белки . Липидный бислой является непроницаемым барьером для большинства водорастворимых молекул и ионов, и их перенос через биомембраны зависит от деятельности транспортных белков. Можно выделить два основных типа этих белков: каналы (поры) и переносчики . Каналы представляют собой туннели, пересекающие мембрану, в которых места связывания транспортируемых веществ доступны на обеих поверхностях мембраны одновременно. Каналы в процессе транспорта веществ не претерпевают каких-либо конформационных изменений, их конформация меняется лишь при открывании и закрывании. Переносчики, наоборот, в процессе переноса веществ через мембрану изменяют свою конформацию. Причем в каждый конкретный момент времени место связывания переносимого вещества в переносчике доступно только на одной поверхности мембраны.
Каналы, в свою очередь, можно разделить на две основные группы: потенциалзависимые и регулируемые химически. Примером потенциалзависимого канала является Na + -канал, его работа регулируется изменением напряжения электрического поля. Иными словами, эти каналы открываются и закрываются в ответ на изменение трансмембранного потенциала . Химически регулируемые каналы
открываются и закрываются в ответ на связывание специфических химических агентов. Например, никотиновый ацетилхолиновый рецептор при связывании с ним нейромедиатора переходит в открытую конформацию и пропускает одновалентные катионы (подрадел 4.7 данной главы). Термины «пора» и «канал» обычно взаимозаменяемы, но под порой чаще понимают неселективные структуры, различающие вещества главным образом по размеру и пропускающие все достаточно малые молекулы. Под каналами чаще понимают ионные каналы. Скорость транспорта через открытый канал достигает 10 6 - 10 8 ионов в секунду.
Переносчики также можно разделить на 2 группы: пассивные и активные. С помощью пассивных переносчиков через мембрану осуществляется транспорт одного типа веществ. Пассивные переносчики задействованы в облегченной диффузии и лишь увеличивают поток вещества, осуществляемый по электрохимическому градиенту (например, перенос глюкозы через мембраны эритроцитов). Активные переносчики транспортируют вещества через мембрану с затратами энергии. Эти транспортные белки накапливают вещества на одной из сторон мембраны, перенося их против электрохимического градиента. Скорость транспорта с помощью переносчиков в очень сильной степени зависит от их типа и колеблется от 30 до 10 5 с -1 . Часто для обозначения отдельных переносчиков используют термины «пермеаза», «транслоказа», которые можно считать синонимами термина «переносчик».
Ферментные функции мембранных белков . В клеточных мембранах функционирует большое количество разнообразных ферментов. Одни из них локализуются в мембране, находя там подходящую среду для превращения гидрофобных соединений, другие благодаря участию мембран располагаются в них в строгой очередности, катализируя последовательные стадии жизненно важных процессов, третьи нуждаются в содействии липидов для стабилизации своей конформации и поддержания активности. В биомембранах обнаружены ферменты - представители всех известных классов. Они могут пронизывать мембрану насквозь, присутствовать в ней в растворенной форме или, являясь периферическими белками, связываться с мембранными поверхностями в ответ на какой-либо сигнал. Можно выделить следующие характерные типы мембранных ферментов:
1) трансмембранные ферменты, катализирующие сопряженные реакции на противоположных сторонах мембраны. Эти ферменты имеют, как правило, несколько активных центров, размещающихся на противоположных сторонах мембраны. Типичными представителями таких ферментов являются компоненты дыхательной цепи или фотосинтетические редокс-центры, катализирующие окислительно-восстановительные процессы, связанные с транспортом электронов и созданием ионных градиентов на мембране;
2) трансмембранные ферменты, участвующие в транспорте веществ. Транспортные белки, сопрягающие перенос вещества с гидролизом АТР, например, обладают каталитической функцией;
3) ферменты, катализирующие превращение связанных с мембраной субстратов. Эти ферменты участвуют в метаболизме мембранных компонентов: фосфолипидов, гликолипидов, стероидов и др.
4) ферменты, участвующие в превращениях водорастворимых субстратов. С помощью мембран, чаще всего в прикрепленном к ним состоянии, ферменты могут концентрироваться в тех областях мембран, где содержание их субстратов наибольшее. Например, ферменты, гидролизующие белки и крахмал, прикрепляются к мембранам микроворсинок кишечника, что способствует увеличению скорости расщепления этих субстратов.
Белки цитоскелета . Цитоскелет представляет собой сложную сеть белковых волокон разного типа и присутствует только в эукариотических клетках. Цитоскелет обеспечивает механическую опору для плазматической мембраны, может определять форму клетки, а также местоположение органелл и их перемещение при митозе. С участием цитоскелета осуществляются также такие важные для клетки процессы, как эндо- и экзоцитоз, фагоцитоз, амебоидное движение. Таким образом, цитоскелет является динамическим каркасом клетки и определяет ее механику.
Цитоскелет формируется из волокон трех типов:
1) микрофиламенты (диаметр ~ 6 нм). Представляют собой нитевидные органеллы - полимеры глобулярного белка актина и других связанных с ним белков;
2) промежуточные филаменты (диаметр 8- 10 нм). Сформированы кератинами и родственными им белками;
3) микротрубочки (диаметр ~ 23 нм) - длинные трубчатые структуры.
Состоят из глобулярного белка тубулина, субъединицы которого формируют полый цилиндр. Длина микротрубочек может достигать нескольких микрометров в цитоплазме клеток и нескольких миллиметров в аксонах нервов.
Перечисленные структуры цитоскелета пронизывают клетку в разных направлениях и тесно связываются с мембраной, прикрепляясь к ней в некоторых точках. Эти участки мембраны играют важную роль в межклеточных контактах, с их помощью клетки могут прикрепляться к субстрату. Они же играют важную роль в трансмембранном распределении липидов и белков в мембранах.


